Polyhydroxyalkanoates (PHA)
A. General background
Biobased polymers
Biobased polymers can be defined as polymeric materials derived from renewable
resources that have been polymerized by chemical and/or biological methods. These
include naturally occurring polymers (e.g., polyhydroxyalkanoate or PHA), modified
natural polymers (e.g., starch and cellulose derivatives) and other bio-chemosynthetic
polymers (e.g., polylactide or PLA, polyvinyl alcohol, polyglycollic acid,
polynucleotides, polyamides and polythioesters). However, it is important to note that not
all biobased polymers are biodegradable. For example, biobased polymers of cellulose
ester derivatives, polythioesters and crystalline PLA are reported to be non-
biodegradable.
Among these biopolymers, PHA has gained major interest due to its excellent
properties over the rest. The properties of PHA resemble some of the properties of
commodity plastics such as polypropylene or low-density polyethylene. In addition, the
complete biodegradability of PHA and utilization of renewable resources for its
production makes it advantageous for numerous applications. The application of
biodegradable polymers like PHA for various purposes can help to reduce the use of
conventional plastic materials, and minimize waste disposal problems.
Polyhydroxyalkanoate (PHA)
PHA is produced by microbial cells as energy reserve material and stored in
dense, protein-covered inclusion bodies termed granules. The building block of PHA is
the (R)-3-hydroxyalkanoic acid monomer unit. The monomers are polymerized into high
molecular weight PHA polymers in the range of 200,000 Da to 3,000,000 Da. The most
common and naturally-occurring microbial PHA is P(3HB).
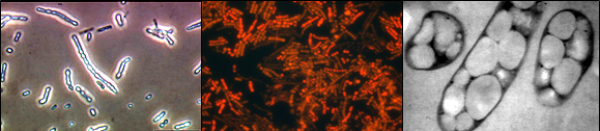
Phase contrast (left), fluorescence (middle) and transmission electron (right) micrographs showing intracellular PHA granules.

Genes involved in PHA biosynthesis
PHA biosynthesis involves the uptake and conversion of carbon sources into precursor molecules that are subsequently polymerized by the enzyme, PHA synthase (PhaC). PhaC is the key enzyme in PHA biosynthesis that catalyzes the polymerization of hydroxyacyl-CoA (HA-CoA) to PHA. Four major classes of PhaC have been distinguished with respect to their primary structures, substrate specificities and subunit composition.
Classes of PHA synthase and the representative species.

Other enzymes involved in the biosynthesis of PHA include the PHA depolymerase (PhaZ), which is responsible for the biodegradation of PHA. Meanwhile, PhaA (β-ketothiolase) and PhaB (NADPH-oxidoreductase), along with PhaC, are encoded by genes that form the phaCAB gene cluster, which plays an important role in the biosynthesis, catabolism and regulation of P(3HB). These proteins catalyze the main reactions in the P(3HB) metabolic pathway. There are also proteins called phasins (PhaP), which are non-enzymatic proteins commonly found at the interface of PHA granules. PhaP influences the quantity and size of granules accumulated by cells.

Organization of genes and enzymes involved in the biosynthesis of PHB in C. necator H16.
PHA biosynthetic pathways
Three main pathways exist for the biosynthesis of PHA. The type of pathway in which PHA is synthesized is dependent on the carbon source provided to the bacterial cells. Pathway I is used by bacteria such as C. necator to produce P(3HB), through the action of three main biosynthetic enzymes (PhaA, PhaB and PhaC). Pathway II and III differ from Pathway I in that fatty acid β-oxidation intermediates and fatty acid biosynthesis intermediates, respectively, are utilized for the production of mcl-(R)-3HA monomers. PhaJ, the (R)-specific enoyl-CoA hydratase generates mcl-3HA-CoA from fatty acid degradation (β-oxidation) pathway while PhaG, the (R)-3-hydroxyacyl-acyl-carrier protein (ACP)-CoA transferase generates mcl-3HA-CoA from the fatty acid biosynthesis pathway. PhaJ and PhaG enzymes act as metabolic links between the fatty acid metabolism to the biosynthesis of PHA.
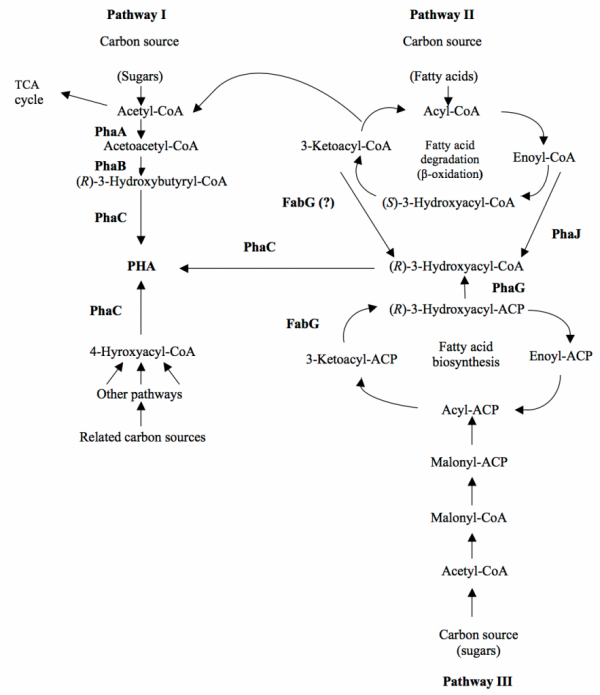
Metabolic pathways that produce various hydroxyalkanoate (HA) monomers for PHA biosynthesis. PhaA, 3-Ketothiolase; PhaB, NADPH-dependent acetoacetyl-CoA reductase; PhaC, PHA synthase; PhaG, 3-hydroxyacyl-ACP-CoA transferase; PhaJ, (R)-specific enoyl-CoA hydratase; FabG, 3-ketoacyl-ACP reductase.
PHA producers and carbon sources
Production of different types of PHA from various carbon sources.

Even though more than 300 different PHA-producing microorganisms have been identified, only a few natural producers are able to produce high concentrations of PHA with high productivity. Strains such as C. necator, D. acidovorans, C. violaceum, Bacilus spp. and some methylotrophs are known for scl-PHA production while P. oleovorans, P. putida and P. aeruginosa can biosynthesize mcl-PHA. Members of the Aeromonas genera (e.g., A. caviae) possess unique and intrinsic abilities to produce scl-mcl-PHA copolymers. Additionally, their robust growth and simple substrate requirements make these bacteria potential hosts for the industrial production of biopolyesters.
The carbon feedstock used in PHA biosythesis is one of the key factors determining the cost of PHA production and the type of PHA produced. The cost of carbon substrate has been estimated to be approximately 28 – 50% of the total production cost. Sugars such as glucose and fructose are among the most common carbon sources; large-scale production of the P(3HB-co-3HV) copolymer has been attempted using a mixture of glucose and propionic acid. Another PHA copolymer, P(3HB-co-4HB), which is an ideal material for biomedical applications, can be produced in the presence of sugars or precursors such as 4-chlorobutyrate, 1,4-butanediol, γ-butyrolactone and 4-hydroxybutyric acid. Meanwhile, plant oils and fatty acid derivatives are relatively cheaper and renewable carbon sources that can generate higher yields of polymer. These oil-based substrates are also suitable for the production of scl-mcl PHA copolymers like P(3HB-co-3HHx), and mcl-PHA using Pseudomonas spp.
References
- Byrom, D. (1992) Production of poly-β-hydroxybutyrate: poly-β-hydroxyvalerate copolymers. FEMS Microbiol Rev 103, 247 – 250.
- Doi, Y. (1990) Microbial polyesters. New York: VCH.
- Khanna, S. and Srivastava, A. K. (2005) Recent advances in microbial polyhydroxyalkanoates. Process Biochem 40(2), 607 – 619.
- Kunioka, M., Kawaguchi, Y. and Doi, Y. (1989) Production of biodegradable copolyesters of 3-hydroxybutyrate and 4-hydroxybutyrate by Alcaligenes eutrophus. Appl Microbiol Biotechnol 30(6), 569 – 573.
- Kunioka, M., Tamaki, A. and Doi, Y. (1989) Crystalline and thermal properties of bacterial copolyesters: Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) and poly(3-hydroxybutyrate-co-4-hydroxybutyrate). Macromolecules 22(2), 694 – 697.
- Lee, S. Y. (1996) Bacterial polyhydroxyalkanoates. Biotechnol Bioeng 49(1), 1 – 14.
- Lee, S. Y., Choi, J. and Wong, H. H. (1999) Recent advances in polyhydroxyalkanoate production by bacterial fermentation: mini-review. Int J Biol Macromol 25(1-3), 31 – 36.
- Madison, L. L. and Huisman, G. W. (1999) Metabolic engineering of poly(3-hydroxyalkanoates): from DNA to plastic. Microbiol Biol Rev 63(1), 21 – 53.
- Rehm, B. H. A. (2003) Polyester synthases: natural catalysts for plastics. Biochem J 376, 15 – 33.
- Sudesh, K., Abe, H. and Doi, Y. (2000) Synthesis, structure and properties of polyhydroxyalkanoates: biological polyesters. Prog Polym Sci 25, 1503 – 1555.
- Sudesh, K. and Iwata, T. (2008) Sustainability of biobased and biodegradable plastics. CLEAN 36, 433 – 442.
- Tsuge, T. (2002) Metabolic improvements and use of inexpensive carbon sources in microbial production of polyhydroxyalkanoates. J Biosci Bioeng 94(6), 579 – 584.
